Research Highlights
Glycan ligands in leukocyte trafficking: Pervading promiscuity
Functional Glycomics (15 June 2007) | doi:10.1038/fg.2007.26Standfirst
Three studies now extend our knowledge about the role of the selectin-glycan interaction in leukocyte migration.
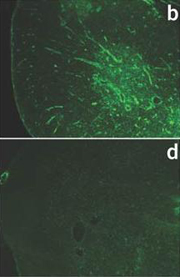
Anti-P-selectin antibody stainings of thymi from wild-type (b) and P-selectin-deficient mice (d)
Selectins and their glycan ligands are present on leukocytes and endothelial cells, and regulate the homing of lymphocytes to secondary lymphoid organs (SLOs) and the trafficking of neutrophils to sites of inflammation. Selectin–glycan interactions allow leukocytes to roll along and tether to the endothelium, where they eventually adhere and enter lymphoid vessels and inflamed tissues. E-, L- and P-selectins are named according to the cell type (endothelial cells, lymphocytes or platelets) on which they were first identified. Their ligands are glycan epitopes – most prominently the tetrasaccharide sialyl-Lewis x – of leukocyte transmembrane glycoproteins such as E-selectin ligand-1 (ESL-1) and the P-selectin glycoprotein ligand-1 (PSGL-1). Three studies now extend our knowledge about the role of the selectin-glycan interaction in leukocyte migration.
A study by Frenette and colleagues in Immunity explores the range of neutrophil ligands that interact with E-selectin on inflamed endothelial cells. Previous work by the authors and other research groups indicated that ESL-1, PSGL-1 and CD44 can all act as E-selectin ligands. An ESL-1 knock down in neutrophils lacking PSGL-1 and CD44 led to a complete loss of E-selectin binding to neutrophils. In vivo, neutrophil rolling velocity was highest and neutrophil recruitment lowest – compared with wild-type neutrophils – when all three ligands were absent. The authors conclude that ESL-1, PSGL-1 and CD44 form the complete set of E-selectin ligands present on neutrophils. Frenette and colleagues propose a model where steady rolling induced by the ESL-1–E-selectin interaction leads to the contact of the entire neutrophil surface to the endothelium. This exposes CD44 for E-selectin binding, which in turn induces CD44 signaling.
But are selectins the only receptors for PSGL-1? And, if not – might PSGL-1 have a function other than supporting leukocyte adhesion? Can altered glycosylation change the ligand binding properties of PSGL-1? The work of Veerman et al., published in Nature Immunology, answers these questions. T cell PSGL-1 interacts with endothelial cell P-selectin to recruit effector T cells to sites of inflammation and regulate homing of naïve T cells to SLOs. Homing also requires T cell chemotaxis elicited by the chemokines CCL19 and CCL21 in the endothelial lumen of lymphatic vessels. Veerman et al. observed that PSGL-1 deficient T cells exhibited reduced migration in response to CCL21 and CCL19 compared to wild-type T cells, and that CCL21 interacted directly with PSGL-1. Activated T cells showed similar chemotactic behavior with or without PSGL-1, suggesting that PSGL-1 does not contribute to chemotaxis after T cell activation. However, when core 2 N-acetylglucosamine transferase 1 expression was ablated in activated T cells, they showed enhanced chemotactic movement in the presence of CCL21, indicating that PSGL-1 glycans are altered by the upregulation of glycosyltransferases occurring during T cell activation. Thus, PSGL-1 contributes to chemotaxis in resting T cells, whereas GlcNAc is transferred to and blocks PSGL-1 in activated T cells.
Guarda et al. report in Nature Immunology that L-selectin expression also varies upon T cell activation. The absence of L-selectin on effector T cells was thought to exclude them from lymph nodes, thus blocking the premature killing of antigen-presenting dendritic cells (DCs). However, Guarda et al. noted that endothelial CXCL9 bound to the T cell receptor CXCR3, leading to the immigration of L-selectin negative T cells to reactive lymph nodes. This might occur as a negative feedback mechanism during infection when a T cell population undergoes clonal expansion. Some of the effector T cells may infiltrate the lymph nodes and stop T cell proliferation by killing the antigen presenting DCs (the proliferation stimulus). It may be interesting to determine whether any of the glycosyltransferases upregulated in activated T cells facilitate the interaction between CXCR3 and CXCL9 as CXCR3 has several potential glycosylation sites.