Featured Articles
Virus hemagglutinins: Sticky structures
Functional Glycomics (13 December 2007) | doi:10.1038/fg.2007.51Standfirst
The receptor binding sites of both the influenza virus B and the measles virus hemagglutinins (HAs) differ strongly from their closest relatives despite a global structural similarity.
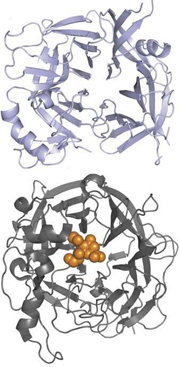
Comparison of measles hemagglutinin structure (top) with human PIV HN structure (bottom) viewed looking directly into the central cavity. Orange, sialic acid bound in the active site of PIV HN. Click here for more details. From Colf et al. 2007.
HAs are glycosylated viral surface lectins that bind to sialic acid molecules on host cell glycans in the first step of host cell virus invasion. Neuraminidase surface proteins on influenza viruses cleave the host cell sialic acids that bind to replicated virus particles, thereby aiding their release from the cell. Instead of possessing distinct proteins, HAs of paramyxoviruses such as measles, parainfluenza (PIV) and mumps viruses possess a HA/neuraminidase (HN) fold. Studies published in the Proceedings of the National Academy of Sciences (PNAS) and Nature Structural and Molecular Biology (NSMB) now pinpoint the different structures for the influenza B/Hong Kong/8/73 virus HA (B/HK HA) and the measles virus HA (MVH).
In PNAS Wang et al. showed that the global structure of HAs from influenza virus A and B is similar despite a sequence similarity of only 20-30%. However, the receptor binding sites of the HAs differed strongly, as documented by a 3.6Å spatial shift between the  carbon atoms of the analogue residues Asp 193 (B/HK HA) and Ser 186 ("Hong Kong" influenza virus A serotype HA (H3 HA)). The authors showed that this is largely due to an F/Y replacement at analogous amino acid residues (95 in B/HK HA and 98 in virus A HAs). An evolutionary advantage of the Y95F exchange in B/HK HA remains to be shown; in contrast to B/HK HA, a Y98F replacement in the H3 HA eliminates sialic acid binding. Interestingly, the B/HK HA glycosylation site at Asn 194 can be lost in chicken egg-grown virus. Wang et al. attribute this loss to the fact that glycans at Asn 194 would interfere with binding of HA to sialic acid molecules being carried by the egg protein allantone. The authors speculate that gain or loss of this glycosylation site may also serve the purpose of modifying influenza B virus antigenicity.
carbon atoms of the analogue residues Asp 193 (B/HK HA) and Ser 186 ("Hong Kong" influenza virus A serotype HA (H3 HA)). The authors showed that this is largely due to an F/Y replacement at analogous amino acid residues (95 in B/HK HA and 98 in virus A HAs). An evolutionary advantage of the Y95F exchange in B/HK HA remains to be shown; in contrast to B/HK HA, a Y98F replacement in the H3 HA eliminates sialic acid binding. Interestingly, the B/HK HA glycosylation site at Asn 194 can be lost in chicken egg-grown virus. Wang et al. attribute this loss to the fact that glycans at Asn 194 would interfere with binding of HA to sialic acid molecules being carried by the egg protein allantone. The authors speculate that gain or loss of this glycosylation site may also serve the purpose of modifying influenza B virus antigenicity.
MVH structure models have been proposed by Colf et al. in NSMB and Hashiguchi et al. in PNAS. Despite the MVH/PIV HN fold similarity, MVH binds to the host cell surface proteins SLAM and CD46 instead of sialic acids, and according to Colf et al., this is related to the marked positional differences of the carbon atoms in the HN fold. Additionally, Hashiguchi et al. point out that the MVH HN fold lack several amino acids that contribute to sialic acid binding in other paramyxoviruses. Furthermore, they show that glycans linked to Asn 200 and Asn 215 make the HN fold inaccessible for sialic acid molecules. The Asn 200-linked N-glycans were found to be located close to each other at the MVH head domain homodimer interface where they occlude a potential sialic acid binding site. Similar to B/HK HA, the MVH glycans are thought to suppress antigenicity and confer conformational variability to the molecule.
These studies all contribute to the understanding of the mode of HA host cell binding and will aid the search for antiviral drugs. They highlight the functional flexibility as well as the structural differences of closely related viruses.