Research Highlights
T cell and fly development: Sugars boost signaling up a Notch
Functional Glycomics (14 February 2008) | doi:10.1038/fg.2008.8Standfirst
Two articles have shown that Drosophila development requires Notch O-glucosylation by the transferase Rumi, and that Notch1 O-fucosylation in the ligand binding domain regulates T cell development in mice.
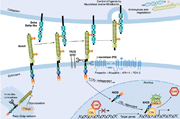
An overview over Notch signaling. From Radtke, F. et al.Notch signaling. EMBO reports 6, 1120-1125 (2005). Please click here for a larger version.
During Notch signaling the intracellular domain of Notch (NICD) is cleaved and translocates to the nucleus where it regulates the expression of Notch target genes. The glycosyltransferase Fringe elongates an initial fucose attached to Notch extracellular domain's (NECD) epidermal growth factor–like (EGF) repeats by the transferase Pofut1. Fringe glycosylation is known to regulate the binding of Delta and Serrate ligands to NECD upon cell–cell contact. In contrast, functional consequences of EGF repeat O-glucosylation are not known yet. Two studies now add a new level to our knowledge of functional glycosylation of Notch.
In Cell, Acar et al. found that developing flies with mutations in the Rumi gene had fewer short sensory bristles (microchaetae) when grown at 25°C than at 18°C. This phenotype was suppressed upon addition of a copy of Notch. These findings suggest that the actions of Rumi are related to the folding or trafficking of Notch. Clonal analyses indicated that Rumi targets the NECD, and a database search pointed to Rumi having glycosyltransferase activity. Indeed, mass spectrometric analysis of the seventh EGF Notch repeat from Rumi-depleted Drosophila S2 cells revealed no change in fucosylation but reduced O-glucosylation, and in vitro assays confirmed that Rumi possesses protein O-glucosylation activity. Acar et al. conclude from their results that Rumi-mediated EGF repeat O-glucosylation is required for Notch function during Drosophila development, and hypothesize that a lack of O-glucosylation affects the folding of Notch which inhibits NICD cleavage and Notch signaling.
In the PNAS, Ge and Stanley analysed mice lacking the O-fucosylation site in Notch1 EGF repeat 12. Notch112f/12f mice were viable but grew more slowly than wild-type mice. T cells from Notch112f/12f mice showed a reduction in NICD cleavage generated by γ-secretase and a reduction in Notch ligand binding. Using flow cytometry, Ge and Stanley showed that Notch112f/12f mice developed fewer double-positive (DP) and single-positive (SP) T cells, and that mutant DP T cells exhibited enhanced apoptosis. Furthermore, the authors demonstrated that the block to T cell development occurs between double-negative (DN) stages 3 and 4 and that the ratio of SP:DP T cells is increased. In sum, a lack of fucosylation in the Notch1 ligand binding domain inhibits the development of early T cell stages and appears to promote the DP-SP transition.
Both studies underscore how Notch glycosylation, initiated by single sugar residues, regulates Notch signaling and influences both Drosophila and mouse development. The results add to the evidence that glycosylation of EGF repeats that are a feature of numerous signaling proteins might be a general way of regulating signal transduction.