Featured Articles
Golgi protein glycosylation: Keeping transferases safe and sound
Functional Glycomics (11 September 2008) | doi:10.1038/fg.2008.40Standfirst
The COSMC chaperone enables the formation of functional T-synthase dimers active in the Golgi apparatus, and most yeast Golgi glycosyltransferases are localized to their compartment by binding to the coat protein I linker Vps74p.
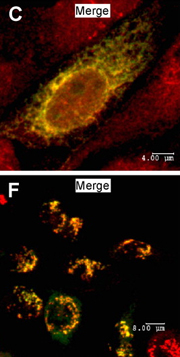
COSMC colocalizes with the ER marker Calnexin (top); T-synthase colocalizes with the Golgi marker mannosidase-II (bottom). © Ju, T. et al. 2008. Originally published in J Cell Biol 182, 531–542 (2008). doi:10.1083/jcb.200711151.
Altered N- and O-glycosylation is a hallmark of many diseases, and Golgi-residing glycosyltransferases perform most glycosylation events. The core 1  3-galactosyltransferase — known as T-synthase as it forms the core 1 O-glycan named T-antigen — has a remarkable feature: it is the only known substrate of COSMC (core 1 3-galactose-transferase-specific molecular chaperone). Binding to COSMC is necessary for forming active COSMC, however, COSMC's effects on T-synthase function are elusive. Apart from this specific transferase, it is unknown which features keep Golgi-resident glycosyltransferases in place. Two studies now enhance our understanding of the factors supporting glycosyltransferase activity in the Golgi apparatus.
3-galactosyltransferase — known as T-synthase as it forms the core 1 O-glycan named T-antigen — has a remarkable feature: it is the only known substrate of COSMC (core 1 3-galactose-transferase-specific molecular chaperone). Binding to COSMC is necessary for forming active COSMC, however, COSMC's effects on T-synthase function are elusive. Apart from this specific transferase, it is unknown which features keep Golgi-resident glycosyltransferases in place. Two studies now enhance our understanding of the factors supporting glycosyltransferase activity in the Golgi apparatus.
After gaining evidence from cell expression studies that human COSMC localizes to the endoplasmic reticulum (ER), Ju et al. investigated how COSMC aids T-synthase function. They transfected the colon-cancer cell line LSC, characterized by absent T-synthase activity and dysfunctional COSMC, with a tagged human T-synthase. After ultracentrifugation and detergent application, the enzyme was recovered from the heavy membrane fraction in a truncated and ubiquitinated form that rapidly underwent proteosomal degradation. These results suggest that the LSC T-synthase resides in the ER lumen where it is fuelled into the ER-associated degradation pathway.
Fully functional T-synthase requires dimer formation. In contrast, Ju et al. recovered most of the T-synthase in LSC cells as non-functional oligomers. Expression of COSMC in LSC cells promoted the formation of T-synthase dimers, which are formed through their association with the COSMC chaperone in the ER. These findings indicate that the interaction of T-synthase monomers with COSMC promotes the dimerization of the T-synthase in the ER lumen, which is subsequently transported into the Golgi apparatus.
Tu et al. noticed that a lack of the peripheral membrane protein Vps74p in yeast cells (vps74 cells) mimicked a lack of Golgi-residing N-glycosyltransferases such as the
cells) mimicked a lack of Golgi-residing N-glycosyltransferases such as the  1,2-mannosyltransferase Kre2p. Using fluorescence-tagged Kre2p, they discovered that the cycling of Kre2p between Golgi apparatus and ER was disrupted in vps74 cells. This observation suggests that the membrane-peripheral Vps74p protein interacts with the cytoplasmic tail of the glycosyltransferase to direct its organelle sorting.
1,2-mannosyltransferase Kre2p. Using fluorescence-tagged Kre2p, they discovered that the cycling of Kre2p between Golgi apparatus and ER was disrupted in vps74 cells. This observation suggests that the membrane-peripheral Vps74p protein interacts with the cytoplasmic tail of the glycosyltransferase to direct its organelle sorting.
Alanine replacement of a semi-conserved motif (F/L)-(L/V)-(S/T), found in the cytoplasmic tails of most yeast Golgi transferases, also resulted in a yeast phenotype similar to a transferase knockout. Finally, the authors discovered that Vps74p acts as a linker, binding both to the cytoplasmic motif of the glycosyltransferases and to COPI (coat protein I) components; furthermore, overexpression of Vps74p led to retention of a high amount of Kre2p in the ER. These results indicate that, instead of a direct binding to COPI components, Vps74p interacts with the cytoplasmic motifs of Golgi-resident glycosyltransferases to sustain their cycling between early and late Golgi stacks and the ER.
Clinical consequences of COSMC mutations have been previously documented. Future work may reveal how alterations to the cytoplasmic tails of Golgi-residing transferases change glycosylation patterns contributing to pathogenesis.