Featured Articles
Golgi: Sugar pHorming
Functional Glycomics (09 July 2009) | doi:10.1038/fg.2009.23Standfirst
Elevated Golgi pH selectively misdirects glycosyltransferase enzymes, impairing complex N-glycosylation.
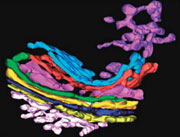
Three-dimensional tomographic reconstruction of the Golgi complex, showing the cis-Golgi network (white) and the stack with the terminal three trans-cisternae in pink, red and cyan. The trans-Golgi network (violet) appears as a tubular network that emerges from the lateral part of the last trans-cisterna (cyan).
To correctly glycosylate and sort proteins and lipids, the Golgi apparatus relies on a highly organized structure and proper acidity, with the luminal pH decreasing gradually from the cis side (near to the ER) to the trans side and post-Golgi secretory compartments. Many cancer cells have insufficiently acidic Golgi bodies which correlate with glycosylation defects. How an abnormal Golgi pH results in abnormal glycosylation has not been clear, partly because most experimental approaches almost completely neutralize the Golgi lumen, simultaneously perturbing the morphology. Sakari Kellokumpu and colleagues used chloroquine to increase the pH more gradually, and discovered that different sialyltransferases can be segregated by their sensitivity to pH-induced mislocalization. Reporting in the Journal of Cellular Physiology, the authors suggest that the purpose of the Golgi pH gradient is to ensure the correct localization of different enzymes involved in complex N-glycosylation.
Core glycan chains are added to proteins in the ER but more complex structures are created by modifications in the Golgi. Sialyltransferase enzymes, which add sialic acid residues to nascent oligosaccharides, include ST3Gal III and ST6Gal I. The former carries out  (2,3)-sialylation of galactose, whereas ST6Gal I sialylates the (2,6) position. When Kellokumpu and colleagues raised the Golgi pH by just 0.2 units, overall morphology and function were preserved. However, although 80% of ST6Gal I maintained its Golgi localization after 24 hours, only 20% of ST3Gal III did, with the remainder re-distributed into vesicle like structures. This mislocalization of ST3Gal III correlated with a reduction in activity: the authors used lectin blotting assays to show that a reporter glycoprotein lacked (2,3)-sialic acid residues, whereas (2,6)-sialylation appeared to be retained.
(2,3)-sialylation of galactose, whereas ST6Gal I sialylates the (2,6) position. When Kellokumpu and colleagues raised the Golgi pH by just 0.2 units, overall morphology and function were preserved. However, although 80% of ST6Gal I maintained its Golgi localization after 24 hours, only 20% of ST3Gal III did, with the remainder re-distributed into vesicle like structures. This mislocalization of ST3Gal III correlated with a reduction in activity: the authors used lectin blotting assays to show that a reporter glycoprotein lacked (2,3)-sialic acid residues, whereas (2,6)-sialylation appeared to be retained.
To investigate which compartment the ST3Gal III was being redirected to, the authors stained cells using antibodies to endosomal markers, and also probed the compartmental pH. Both approaches indicated a late endosomal/lysosomal location. However, proteins can be trafficked directly to late endosomes from the Golgi, or alternatively sent first to the plasma membrane and then re-internalized, so the team investigated which route was followed. They used cell surface antibody binding to find that the redirected ST3Gal III did reach the plasma membrane, indicating that it escaped the Golgi via bulk flow to the cell surface. Whether this escape results from improper retention in the Golgi, or from improper retrieval of the enzyme from maturing compartments back to earlier compartments, depends on which of these two presently competing models for Golgi transport eventually proves correct.
Why do ST3Gal III and ST6Gal I differ in their sensitivity to pH? Both enzymes normally reside in the Golgi stacks, the authors found, ruling out separate initial sub-compartmentalization. That ST6Gal I, unlike ST3Gal III, forms dimers was found to be irrelevant. Domain swapping experiments implicated the luminal domain of ST6Gal I in maintaining Golgi localization despite rising pH, but any potential interactors remain to be identified. ST3Gal III is probably not the only terminal glycosylation enzyme susceptible to mislocalization by a moderate increase in pH: the changed size of a reporter glycoprotein in the chloroquine-treated cells suggests that (2,3)-sialic acid was not the only glycan lost.
Further work is needed to show how the Golgi pH gradient affects enzyme localization. Improved understanding might provide a means of modulating special glycosylation steps during protein production, and will shed light on glycosylation defects in cancer.