Featured Articles
Carbohydrate length: How to say when
Functional Glycomics (09 July 2009) | doi:10.1038/fg.2009.24Standfirst
Binding of carbohydrate polymerase to both ends of an initiator allows it to control product length without a template.
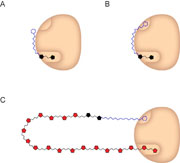
Proposed tethering model for GlfT2 (tan). (A) With acceptors that cannot occupy the lipid-anchoring site, only short oligomers are generated. (B) When GlfT2 interacts with acceptors that can bind to both sites, longer oligomers are obtained. (C) When the polymer chain becomes long, polymer dissociation competes with further elongation. PNAS (Published online 2 July 2009) doi: 10.1073/pnas.0901407106 Artist: Adam Steinberg.
Polysaccharides have diverse cellular functions and come in a range of lengths. Whereas large structural carbohydrates such as cellulose contain thousands of sugar residues, and signaling polysaccharides often have hundreds, oligosaccharide linkers may be only tens of units long.
Unlike DNA polymerases, carbohydrate polymerases have no template to follow and little is known about how these enzymes achieve length control. Laura Kiessling and colleagues used synthetic substrates to probe the mechanism of a mycobacterial glycosyltransferase. Reporting in Proceedings of the National Academy of Sciences of the USA, they propose that this enzyme has two binding sites for its substrate, one for either end of the chain. Once long enough, the increasingly unruly chain is more likely to dissociate from the enzyme, producing products within the desired length range.
There are two ways that a polymerase may elongate its substrate: either bind and release it in multiple rounds, adding a single repeat unit each time (distributive polymerization) or keep hold of it through multiple rounds (processive polymerization). Distributive polymerization results in a statistical Poisson distribution of product lengths, whereas the lengths of processive polymerization products depend on when they are released from the enzyme and the distribution is skewed towards the desired length. Without a template, it was unknown what the trigger for processive carbohydrate polymerase release might be. One such enzyme is galactofuranosyltransferase (GlfT2), which produces a mycobacterial cell-wall lipid-protein connector termed galactan. GlfT2 makes galactan by adding galactofuranosyl (Galf) residues to a lipid-linked oligosaccharide acceptor. Although in vitro experiments had shown that GlfT2 can add multiple Galf units to the same acceptor, the length of these products did not approach the 20–40 seen in vivo, and it was unknown how regulated galactan length was achieved.
To investigate length control by GlfT2, Kiessling and colleagues synthesized acceptor molecules of Galf joined to lipids of varied lengths. After allowing polymerization for set time periods, products were analyzed by mass spectrometry. Using acceptor molecules that were designed to be similar to the natural lipid-linked substrate, the authors first showed that with the right conditions GlfT2 alone is able to produce products within the desired 20–40 unit length range. The distribution of product lengths favored processive polymerization by the enzyme.
Other polymerases, including DNA polymerase, have an extended binding site that interacts with repeating units of growing product, in effect monitoring the length, and it has been proposed that such a site might regulate product length for template-independent polymerases such as GlfT2. However, the disaccharide component of the acceptor molecules was not sufficient to produce extended polymers: the authors found that the nature of the lipid substituent is key in that it must be long enough. This suggested the presence of a secondary lipid binding site on the enzyme, in addition to the active site where the glycan binds. Kinetic studies of dissociation rates supported multivalent binding of the substrate by GlfT2. Furthermore, whereas acceptors with short lipids were unable to produce extended polymers, increasing the lipid length also increased the length of the polymer produced.
The strength of binding between enzyme and product, or 'tethering' of the product, effects how quickly any enzyme releases its product and terminates a reaction. By binding both ends of the growing product, GlfT2 may tether the molecule sufficiently to prevent premature release. Increased movement within a lengthening chain would then make dissociation increasingly likely, limiting GlfT2 products to their specific length range.