Research Highlights
N-linked glycosylation: Reducing options
Functional Glycomics (09 July 2009) | doi:10.1038/fg.2009.25Standfirst
An oligosyltransferase subunit regulates substrate recognition and conformation with a redox-sensitive loop.
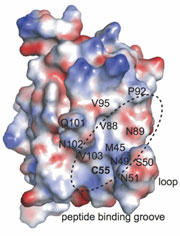
Surface representation of oxidized Ost6p showing the proposed peptide binding groove, colored by electrostatic surface potential (red, acidic; white, neutral; blue, basic). PNAS (Published online 23 June 2009) doi: 10.1073/pnas.0812515106
Oligosaccharyltransferase (OTase) catalyzes the shift of a saccharide from a lipid scaffold to specific asparagines of selected protein substrates as part of the protein modification pathway in the endoplasmic reticulum. Further modifications to this initial N-linked glycan influence protein folding, trafficking and function, and are integral to a wide variety of biological processes, yet the functional details of the OTase complex are unclear. The yeast OTase subunits Ost3p and its paralog Ost6p are known to influence substrate selection, but the basis for this specificity is unknown. Now Schulz et al. provide evidence that these proteins are functional oxidoreductases and contain a redox-dependent substrate binding site. Reporting in Proceedings of the National Academy of Science of the USA, the authors integrate this information to postulate a new model of transient disulfide bonds driving Ost3/6p function within the OTase machinery.
The first clue to the function of Ost3/6p came from the presence of a CxxC motif that is characteristic of thioredoxin-like proteins. These enzymes are capable of forming, reducing, and isomerizing disulfide bonds. With this insight, Schulz et al. speculated that Ost3p and Ost6p could be oxidoreductases. To test this hypothesis, the authors mutated the cysteine residues in the thioredoxin motif to serines, and observed changes in glycosylation for a subset of OTase substrates. In vitro assays with peptide and protein substrates indicated that Ost6p was a functional oxidoreductase (reducing disulfide bonds) but not an isomerase. Finally, the redox potential of the protein was found to be -275 mV, which suggests that the CxxC motif of Ost6p is oxidized to form a disulfide bond in vivo.
The authors turned next to the structure of these enzymes. Three crystal structures of Ost6p at different oxidation states highlighted a distinctive loop near to the CxxC motif. The loop is conserved throughout Ost3/6p sequences and the authors noted that, when the protein was oxidized, it formed part of a potential peptide binding groove. In contrast, in the reduced state, the loop became flexible, disrupting this binding site. This correlated with peptide binding assays, which showed that some model substrates were only bound by the oxidized protein and not the reduced form.
Combining this information led the authors to speculate that the disulfide bond of Ost6p (and, by analogy, Ost3p) serves as a transient, covalent anchor for the nucleophilic, reduced cysteines of incoming unfolded peptide sequences. The attachment of these sequences, coupled with additional binding energy gained from interactions in the peptide binding groove, prevent the oxidative folding of the protein and positions these glycosylation substrates next to the catalytic OTase subunit long enough to facilitate glycan transfer. Release of this intermolecular disulfide bond would restore the oxidation state of the OTase subunit and would allow the glycosylated protein substrate to continue the folding pathway.
The assembly and transfer of complex oligosaccharides by OTase is a critical step in the maturation of protein sequences. Investigating the role of individual subunits within this complex machinery will help to delineate sources of substrate specificity and regulation. This study highlights a redox-dependent structural rearrangement as a surprising new feature of thioredoxin-like function, and improves our understanding of OTase activity by establishing the dual function of Ost3/6p in selecting and tethering glycosylation substrates.